Amir Gharib1 ![]() ,
Zohreh Faezizadeh1,
Masoud Godarzee2
,
Zohreh Faezizadeh1,
Masoud Godarzee2
For correspondence:- Amir Gharib Email: amirgharib@gmail.com Tel:+986623500201
Received: 18 March 2016 Accepted: 17 July 2016 Published: 30 August 2016
Citation: Gharib A, Faezizadeh Z, Godarzee M. Inhibition of telomerase activity and cell growth by free and nanoliposomal forms of punicalagin in human leukemia cell line K562. Trop J Pharm Res 2016; 15(8):1621-1630 doi: 10.4314/tjpr.v15i8.4
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To prepare punicalagin-loaded nanoliposome and compare its anti-telomerase activity in K562 cell line with that of free punicalagin.
Methods: Punicalagin-loaded nanoliposomes were prepared by extrusion method, and the efficiency of punicalagin entrapment was determined by high-performance liquid chromatography (HPLC) method. The anti-proliferation effect of the punicalagin in the free and nanoliposomal forms at various doses (0 - 100 µg/mL) and times (0 - 72 h) on K562 cell line was investigated using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide ((MTT) assay. Changes in telomerase activity, following treatment with punicalagin, of the free and nanoliposomal forms were measured by telomeric repeat amplification protocol-enzyme-linked immunosorbent assay.
Results: The entrapment efficiency of punicalagin was 86.7 ± 1 %. Treatment of K562 cells with punicalagin, particularly in the nanoliposomal form, resulted in significant induction of apoptosis and inhibition of cell growth. Furthermore, the telomerase activity of the nanoliposomal punicalagin-treated cells was significantly inhibited in a time- and dose-dependent manner.
Conclusion: Punicalagin shows a novel mechanism of anti-telomerase activity, particularly in the nanoliposomal form, and may provide a basis for the future development of anti-cancer therapy.
Introduction
Telomerase is a reverse transcriptase enzyme responsible for the renewal of chromosomal ends, the so-called telomeres [1]. Increase in telomerase activity is closely related to expression of subunit human telomerase reverse transcriptase (hTERT) which is widely expressed in almost all tumor-derived human cell lines, and present in at least 85 % of human tumor samples [2]. Previous studies have showed that expression of hTERT and activation of telomerase is a crucial step in tumorigenesis [3]. It has been reported that inhibition of telomerase triggers apoptotic cell death in various cancer cell types [4].
In cancer therapy research, many different synthetic chemical agents have been proposed for telomerase inhibition but most of these compounds have severe toxic side effects on normal cells [5]. As a result, extensive studies have been carried out in search of compounds that are capable of inhibiting telomerase activity and retarding growth of cancer cells without affecting normal cells [6,7]. Many studies have shown that certain plant-derived compounds have this property and were able to inhibit telomerase activity in several cancer cell lines without affecting normal cells [8,9].
Punicalagin () a phenolic compound is found in some plants such as Punica granatum, Terminalia catappa and Combretum molle [10]. It has inhibitory effect on different kinds of cancer cells including prostate and colon cancer cells [11,12]. However, the in vivo instability of punicalagin has restricted its use in biomedical research [13]. Previous studies showed that encapsulation of plant-derived materials in nanocarriers such as nanoliposomes markedly reduced their in vivo instability and increased their effectiveness [14,15].
A liposome is a colloidal and spherical vesicle that can be used as a vehicle for the administration of pharmaceutical drugs and nutrients [16]. Liposomes have diameter ranging from a few nanometers to several micrometers, and compose of natural phospholipids and other lipids, such as cholesterol [14,15]. To date, the anti-proliferative and anti-telomerase activities of punicalagin in the free and nanoliposomal forms against human K562 cell line have not yet been studied. The aim of the present study was to prepare punicalagin-loaded nanoliposomes and investigate their in vitro antiproliferative and anti-telomerase activities against human leukemia cell line K562.
Methods
Materials
Fetal bovine serum (FBS), RPMI-1640, trypan blue, streptomycin and penicillin were purchased from Gibco BRL (Gaithersburg, MD, USA). Punicalagin (purity ≥ 98 %), soy lecithin, cholesterol and Hoechst 33342 were purchased from Sigma (St. Louis, MO, USA). TeloTAGGG telomerase polymerase chain reaction (PCR)-enzyme-linked immunosorbent assay (ELISA) Plus kit and Annexin V-propidium iodide (PI) apoptosis Detection Kit were obtained from Roche (Applied Science, Germany). Acetic acid and mwere purchased from Merck (Darmstadt, Germany).
Preparation of punicalagin-loaded nanoli-posomes
Punicalagin-loaded nanoliposomes were prepared by extrusion method, as described previously with some modifications [16]. Briefly, soy lecithin and cholesterol (5:1 molar ratio) were dissolved in chloroform and then dried to a lipid film with a rotary evaporator (Brinkman) at 30 °C under controlled vacuum. The dried lipids were dispersed by agitation in 1 mL of punicalagin solution (100 μg/mL) and sonicated at 4 °C in ultrasonic bath (Braun-sonic 2000).
Thereafter, punicalagin-loaded nanoliposomes were obtained by extruding of nanoliposomal suspension 10 times through a polycarbonate membrane filter with 100 nm pores. Excess free punicalagin and larger lipid aggregation were separated by ultracentrifugation (100,000 g, 30 min). The control nanoliposomes were prepared similarly, but PBS (pH 7.4) was used instead of the punicalagin solution.
Characterization of nanoliposomes
Punicalagin content of the nanoliposomes was determined by HPLC method [17] following dissolution in 0.1 % Triton X-100. After dissolution, 20 µL of lysate was injected into the HPLC column (C18, 4.5 × 150 mm, 5 µm, Phenomenex, Torrance, USA). The diode array UV detector and the mobile phases, including water (A); 88 % water + 12 % methanol (B); 20 % water + 80 % methanol (C); methanol (D) was used. The elution started with 100 % A (5 min) and a gradient was used to obtain 100 % B at 10 min, holding it isocratic for 3 more minutes and from 13 to 35 min. Subsequently, a linear gradient was used to obtain 50 % B and 50 % C, and then 100 % C at 40 min and the column was washed with 100 % D at 42 min. The flow rate was 0.9 mL/min. Finally, the loading (L) of punicalagin was calculated as in Eq 1.
L (%) = {(WnV)/(VWi)}100 ………………. (1)
where Wn is the amount of punicalagin in nanoliposomes, V the total sample volume tested and Wi the initial amount of punicalagin used in preparing the nanoliposomal formulation. The polydispersity index, size distribution, zeta potential and mean particle size of the nanoliposomes was evaluated using a Malvern zetasizer (Malvern Instrument, Worcestershire, UK) apparatus, as reported previously [8]. The analysis of the size and shape of the punicalagin-loaded nanoliposomes were carried out by cryo-TEM, as previously described [18].
Cell culture
The human chronic myelogenic leukemia cell line K562 was purchased from Pasteur Institute of Iran (Tehran, Iran). The K562 cells were maintained in RPMI-1640 medium supplemented with 10 % FBS along with streptomycin (100 µg/ml) and penicillin (100 units/ml) and were grown in a humidified incubator (37 ºC and 5 % CO2). Punicalagin was dissolved in the deionized water to obtain a 100 µg/ml stock solution and all subsequent dilutions were made in RPMI medium.
Cell proliferation assay
Cell proliferation was determined using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay as previously described [19]. In brief, 5 × 103 cells were incubated in 96 well plates in the presence of different concentrations of punicalagin in the nanoliposomal and free forms (0, 10, 50 and 100 µg/mL) for 24, 48, and 72 h in a final volume of 200 µl. At the end of the treatment, 20 µL of MTT (5 mg/mL in PBS) was added to each well and then incubated at 37 °C for an additional 2 h. The resultant precipitate was dissolved in 200 µL of DMSO and the viability of cells was assessed by measuring the optical density at 570 nm. Three replicates were done for each concentration of punicalagin in the free and nanoliposomal forms.
Nuclear staining with Hoechst 33342
K562 cells were incubated with punicalagin in the free and nanoliposomal forms (0, 10, 50 and 100 µg/mL) for 72 h and washed with PBS buffer. After fixation in freshly prepared ice-cold paraformaldehyde (0.1 %) for 10 min, the cells were stained with Hoechst 33342 (50 µg/mL) for 1 min in the dark and morphologic changes were observed under the fluorescent microscope. Thereafter, the percentage of apoptotic cells was assessed after counting at least 100 cells per treatment group.
Flow cytometric analysis
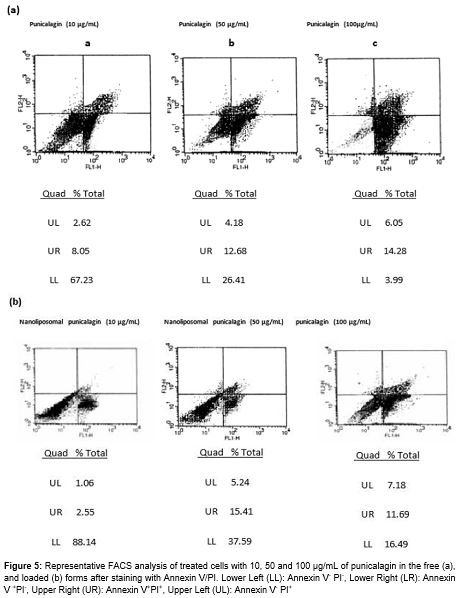
The K562 cells were treated with varying concentrations of the punicalagin and its nanoliposomal form (0, 10, 50 and 100 µg/mL) for 72 h. At the end of each treatment, the cells were collected and quantitative apoptotic death assay was done by Annexin V and PI staining following the manufacturer’s protocol. Apoptotic cells were measured immediately by flow cytometry.
Telomerase activity assay
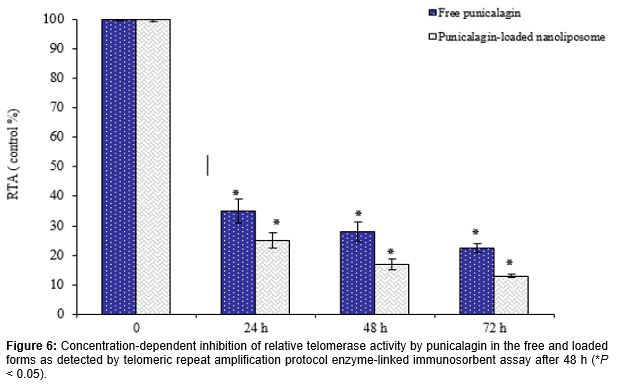
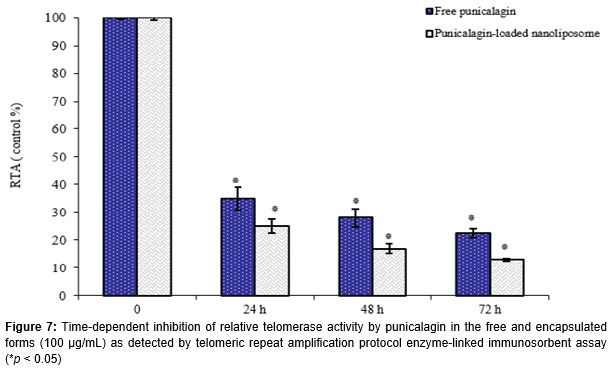
The K562 cells were first treated with different concentrations (0, 10, 50 and 100 µg/mL) of the punicalagin in the encapsulated and free forms for 72 h. Moreover, in order to investigate the effect of different incubation times on telomerase activity, punicalagin in the free and encapsulated forms at 100 µg/mL was incubated for different time intervals (0, 24, 48 and 72 h). At the end of each treatment, the cells were harvested and washed twice with washing buffer and lysed in the lysate buffer. The protein concentration was determined using Bradford method [20]. The telomerase activity was assessed using TeloTAGGG telomerase PCR-ELISA plus Detection Kit according to the manufacturer’s protocol (Roche Applied Science, Germany). In the presence of biotin-labeled primers cell extracts were incubated for 30 min and the telomeric repeats were built by cell-extracted telomerase.
Thereafter, the elongated products as well as a special internal standard were amplified by PCR. The PCR products were denatured and split in two aliquots which were separately hybridized to digoxigenin-labeled detection specific probes and allowed to bind to a streptavidin coated, 96-well plate. Then the biotin-labeled PCR products were detected using peroxidase-conjugated antibody. Absorbance of the developed blue color was measured at 450 nm by STAT-FAX 3200 ELISA reader (Awareness Technology, Inc., USA). As the negative control, each extract was heated for 10 min at 95 °C prior to the PCR step. Thereafter, relative telomerase activity (RTA) of each sample was determined according to the instruction of TeloTAGGG Telomerase PCR-ELISA PLUS kit.
Statistical analysis
The results are presented as mean ± standard deviation (SD). Analysis of variance was done to determine significant level among the test groups and differences with p < 0.05 were considered statistically significant. Data were analyzed using SPSS software (version 19.0, SPSS, Inc., IL, USA).
Results
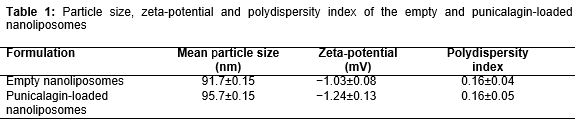
The data revealed that percentage of punicalagin entrapment efficiency was 86.7 ± 1 %. shows the mean particle size, zeta-potential, and polydispersity index of empty and punicalagin-loaded nanoliposomes. The size homogeneity of empty and loaded nanoliposomes suggested that punicalagin was entrapped into nanoliposomes, according to previous reports [14-16]. In addition, zeta-potential and polydispersity index of the punicalagin-loaded nanoliposomes showed that nanoliposomes have appropriate stability in the aqueous dispersion [14].
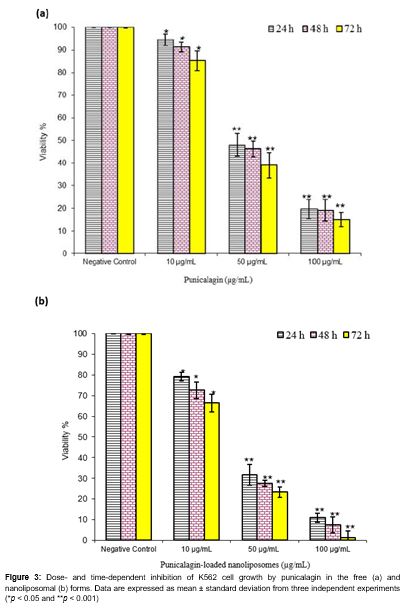
The viability of the K562 cells after treatment with punicalagin in the free and encapsulated forms was examined by MTT assay. The results revealed that the cell proliferation was inhibited in the K562 cells in a time- and dose-dependent manner (). In all conditions, the punicalagin-loaded nanoliposomes were more effective than those of free punicalagin on K562 cell growth. As shown in , the extent of inhibition increased significantly at 24 h with the lowest concentration of punicalagin in the encapsulated and free forms which continued to rise at 48 and 72 h durations at the maximum concentration.
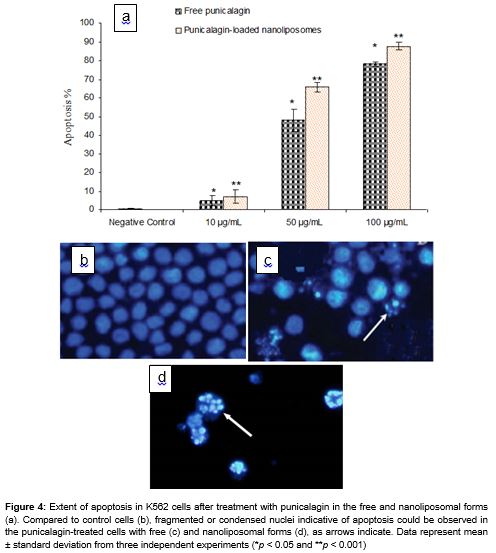
Cryo-TEM analysis revealed that nanoliposomes had a fine homogeneity and were spherical in shape (). Under a fluorescence microscope, punicalagin-treated cells stained after 72 h showed characteristic apoptotic features such as chromatin condensation and nuclear fragmentation (c and 4d). In particular, in the presence of punicalagin-loaded nanoliposomes, the percentage of the apoptotic cells was increased in a dose-dependent manner and percentage of apoptotic cells was also higher than in those exposed to free punicalagin (a).
Flow cytometry results
As shown in the representative FACS analysis scatter-grams, cells treated with punicalagin, particularly in the encapsulated form at 10, 50 and 100 µg/mL for 72 h showed strong shift from viable cells to the early and late apoptotic cells ().
When compared to untreated cells, the RTA of K562 cells were significantly suppressed in a concentration and time-dependent manner after treatment with punicalagin, particularly in the loaded form (Figures 6 and 7). Compared to control group, results showed that RTA of cells treated with 100 µg/mL punicalagin in the free and nanoliposomal forms at 72 h was reduced to 22.5 ± 1.5 and 13.1 ± 0.5 %, respectively (). Similarly, as shown in , RTA of K562 cells treated with punicalagin in the free and loaded forms was reduced to 28.2 ± 1.2 % and 17.1% ± 1.8 at 48 h respectively, as compared to control group (p < 0.05).
Discussion
Punicalagin is an ellagitannin, found in Pomegranate (Punica granatum L.) and some other plants. [10]. Evidence of the health benefit of punicalagin are manifested in its strong antioxidative, anti-cancer, anti-inflammatory, and anti-atherosclerotic activities [11,12]. Punicalagin is a major antioxidant polyphenol in pomegranate juice, where it may reach levels of up to 1 g/L of juice [21]. It has potential antiviral effect against enterovirus 71 and could be used for the therapy of various immune pathologies [22].
Moreover, punicalagin has an inhibitory effect on some cancer cells, including HT-29 and HCT116 colon cancer cells [23]. It was reported that telomerase activity was an important marker for tumorigenesis and inhibition of this enzyme could be used for the development of anti-proliferative agents and anti-tumor drugs [1-4].
Plant-derived compounds that can suppress telomerase activity might be potential candidates for cancer therapy because telomerase activity is found in a wide variety of cancer cells, including leukemia; this phenomenon could enhance resistance to apoptosis through multiple mechanisms [8,9,15].
However, the main problem associated with the application of some plant-derived compounds is biological instability [13]. Previous studies showed that the incorporation of plant-derived compounds into nanoliposomes could overcome this limitation [14]. It has been reported that encapsulation of plant-derived materials in nano-liposomes can increase their stability and effectiveness [24].
In this study we found that punicalagin, in particular in the loaded form, significantly inhibited proliferation and telomerase activity in K562 cells in a concentration as well as time dependent manner. The data also indicated that anti-telomerase activity of punicalagin-loaded nanoliposomes in K562 cells was higher than those of free punicalagin.
Several hypotheses, including increased penetration of plant-derived compounds into cells and stability of encapsulated materials may explain the mechanism of enhanced anticancer efficacies of this nanoliposomal formulation [25-30]. Our data showed a significant and positive correlation between telomerase inhibition and induction of apoptosis.
Similar anticancer activity had been reported in previous investigations with other liposomal natural plant products such as celastrol [25], epigallocatechin-3-gallate [24], gossypol [26], wagonin [27], berberin [28], curcumin [29] and tea polyphenol [30] in various cancer cell lines. Therefore, punicalagin and other effective plant-derived agents in the free and encapsulated forms may be considered as promising strategy to develop new anticancer drugs.
Conclusion
Nanoliposomal punicalagin with acceptable characteristics has been successfully prepared. The findings of this study demonstrate that punicalagin, in the nanoliposomal form has potent anti-telomerase effect on K562 cells, and also time- and concentration-dependently inhibits cell growth in this cell line. Therefore, the developed nanoliposomal formulation may provide a basis for the future development of anti-telomerase agents for leukemia therapy.
Declarations
Acknowledgement
References
Archives


News Updates