Morteza Hadizadeh1,
Amin Norouzi2,
Rohollah Taghadosi3,
Samane Mohebi2,
Mohsen Mohammadi4,
Amir Hasanzadeh5,
Majid Taati Moghadam2 ![]()
For correspondence:- Majid Moghadam Email: majidtaati1367@yahoo.com Tel:989119343489
Received: 6 July 2016 Accepted: 12 December 2016 Published: 31 January 2017
Citation: Hadizadeh M, Norouzi A, Taghadosi R, Mohebi S, Mohammadi M, Hasanzadeh A, et al. Prevalence of qnr, intI, and intII genes in extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli isolated from clinical samples in Iran. Trop J Pharm Res 2017; 16(1):141-147 doi: 10.4314/tjpr.v16i1.18
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the prevalence of qnr, intI, and intII genes in extended spectrum beta-lactamase (ESBL)-producing Escherichia coli isolated from clinical samples in Kerman, Iran.
Methods: A total of 127 E. coli were collected from clinical samples in Kerman hospitals. The antibiotic susceptibility test was performed using disc diffusion method, while the presence of ESBL-producing E. coli was determined by phenotypic confirmatory test. Furthermore, the presence of qnrA, qnrB, qnrS, intI, intII, and β-lactamase-encoding genes was detected by polymerase chain reaction (PCR). Finally, the data were analyzed and associations between different genes and antibiotic resistance were evaluated.
Results: The highest and lowest rates of resistance were observed against ampicillin (72.4 %) and imipenem (2.3 %), respectively. Also, 41.7 % of the isolates produced ESBL-enzymes. The qnrS and genes were detected in 6.3 and 0.78 %, respectively, of the isolates, while qnrA gene was not detected in the current study. The results revealed that 64.5 and 10.2 % of isolates carried intI and intII genes, respectively. Data analysis showed a significant association between ESBL production and class I integrin gene in E. coli isolates.
Conclusion: Due to the variation in the resistance patterns of E. coli against antibiotics in different geographical regions, antimicrobial treatments should be based on local experience. Also, the coexistence of ESBL and intI gene in the majority of E. coli isolates suggests that care should be taken in choosing antibiotic therapy.
Introduction
Extended-spectrum β-lactamase (ESBLs) such as E. coli and Klebsiella pneumonia are frequently prevalent in the Enterobacteriaceae family [1,2]. In recent years, ESBL-producing E. coli has mostly been implicated in nosocomial infections [3]. ESBL-producing organisms are resistant not only to beta-lactams, but also to most other antibiotic families such as aminoglycosides and quinolones, and that results in the limitation of therapeutic options [3,4]. ESBLs are enzymes commonly derived from TEM or SHV families of enzymes [4]. CTXM is also a concern and has become increasingly prevalent in the world since 1995 [5]. The emergence of quinolone resistance among human gram-negative pathogens is also of concern [6,7].
Several mechanisms contribute to the resistance to quinolone antibiotics; plasmid-mediated quinolone resistance qnr genes are a common mechanism [8]. Although the qnr gene is associated only with low-level resistance to fluoroquinolones, the presence of the qnr gene along with chromosomal mutations can contribute to high levels of resistance. It has also been suggested that the relationship between qnr and ESBLs is co-associated with ESBL-encoding genes. High levels of quinolone resistance in clinical strains of E. coli and other ESBL-producing bacteria have been reported in various studies [9-11]. However, the majority of recent studies on ESBL genes found that ESBL genes were located on integrons. Therefore, ESBLs can be located on integron genes and easily transferred into various bacteria [12].
The prevalence of antibacterial resistance among E. coli species is an important problem. Antimicrobial resistance patterns are different in qnr and ESBL-producing species [6,7,13]. The main purpose of the current study was to detect the relationships between ESBL and qnr and between ESBL and integrons (intI and intII) to demonstrate that ESBLs are located on integrons or plasmids.
Methods
Ethical issues
This work was approved by the ethical committee of Department of Infectious Diseases, School of Medicine, Kerman University of Medical Sciences, Kerman, Iran (ref no. 1394.223). Written informed consent was obtained from the patients or a close relative. Written informed consent was obtained from the patients or their families. All stages of the studies pertaining to patients to the patients followed international guidelines for ethics and research [14].
Bacterial isolates
In the current study, 127 E. coli from urine, blood, wound, and other bodily fluids were collected from Kerman hospitals. All 127 E. coli were identified biochemically using standard methods including motility, indole production, negative citrate test, glucose, sucrose, and lactose fermentation in TSI agar, and positive MR reaction in MRVP medium. After E. coli was identified, the isolates were evaluated for antimicrobial resistance.
Antimicrobial susceptibility test and β-lactamase determination
Antibiotic susceptibility tests were carried out using the disc diffusion method according to Clinical and Laboratory Standard Institute guidelines (CLSI). Antibiotic discs including ampicillin, ceftriaxone, cefotaxime, ceftazidime, co-trimoxazole, nalidixic acid, ciprofloxacin, gentamicin, amikacin, and imipenem were used. ESBL-producing E. coli was determined using the phenotypic confirmatory test. The phenotypic test was performed using cefotaxime-clavulanic acid or ceftazidime-clavulanic acid as two-disk synergism versus ceftazidime or cefotaxime alone. If the inhibition zone around the synergism disks compared with ceftazidime or cefotaxime alone was ≥ 5mm, then these species was found to be ESBL-producing. E. coli (ATCC 25922) was used as a negative control to determine antibiotic susceptibility.
Detection of qnr, intI, intII, and β-lactamase-encoding genes
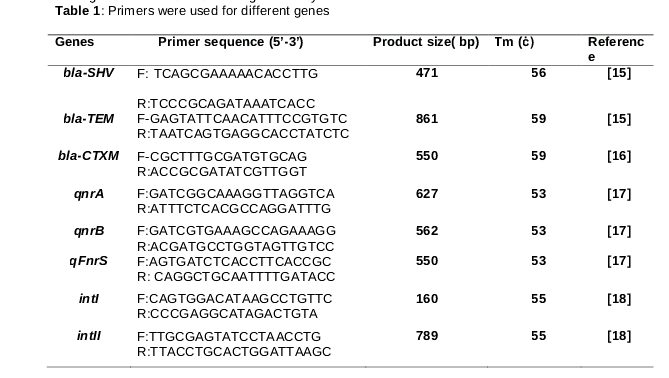
The presence of intI, intII, qnrA, qnrB, qnrS, bla-TEM, bla-SHV, and bla-CTXM genes was detected by standard PCR amplification. First, E. coli colonies were grown in MacConkey agar and then transferred into 1 ml of water in an Eppendorf tube to be boiled. After 15 mins boiling, DNA templates were extracted for PCR amplification. PCR reactions were performed in a final volume of 25 µL using 3 µL of DNA solution, 12.5 µL Qiagen HotStar Taq polymerase Master Mix, 10 pmol primer, and water. PCR cycling conditions were as follows: denaturation step at 94 oC for 5 min, followed by 30 cycles of denaturation at 94 oC for 1 min, extension at 72 for 40 s, and the final extension step at 72 oC for 5 min. All primers used in the current study are listed in . PCR products were detected using electrophoresis on a 1 % agarose gel. The agarose gel was stained with ethidium bromide. The agarose gel was illuminated by UV and then analyzed. Figures 1 and 2 demonstrate the locations of the genes on the agarose gel. Klebsiella pneumonia (ATCC 700603) was used as the positive control for bla-SHV and bla-CTXM genes. E. coli (ATCC 35218) was also used as the positive control for the bla-TEM gene, and E. coli (ATCC 25922) was considered as the negative control. Acinetobacter baumannii (TMU1, TMU2) were used as the positive controls for integron classes I and II. Furthermore, E. coli J53 strains containing pMG252, pMG298, and pMG306 were used as qnrA, qnrB, and qnrS-positive controls, respectively.
Statistical analysis
Data was analyzed using SPSS, version 22 and is presented as mean ± standard deviation (SD). Associations between different genes and antibiotic resistance were also evaluated using the chi-square (X2) test. P < 0.05 was considered statistically significant.
Results
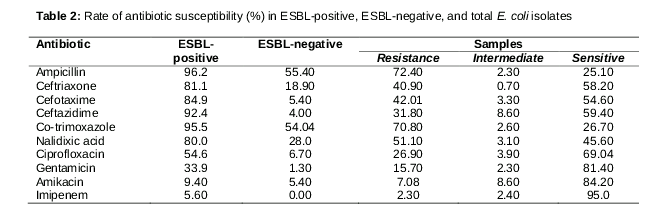
The findings indicate that the resistance rates of ESBL-producing isolates to all antibiotics were higher than those of non-ESBL-producing isolates. In the current study, 127 E. coli were isolated from a range of clinical specimens including the bodily fluids of patients with urinary tract, blood, wound, and other types of infection. E. coli isolates were most frequently isolated from patients with urinary tract infection (89.7 %); the isolated samples of blood, wound, and bronchial fluid were 3 (2.3 %), 6 (4.7 %), and 5 (3.9 %), respectively. Of all 127 samples, 82 (64.5 %) were isolated from females compared to 45 (35.4 %) samples isolated from males. As shown in , even though imipenem was the most effective antibiotic against E. coli isolates, susceptibility to amikacin, ciprofloxacin, and gentamicin was also high. Ninety-two isolates (72.4 %) were resistant to ampicillin, a higher percentage than for any other antibiotic. Resistance to co-trimoxazole and nalidixic acid was also significant. E. coli clinical isolates showed variable susceptibility to different antibiotics. For example, isolates recovered from urinary tract infections were more resistant to ciprofloxacin and gentamicin than others, but resistance to imipenem, ampicillin, and ceftriaxone was higher in wound and bronchial fluid samples. Details of the antibiotic susceptibility tests are presented in .
The results of this study further showed that various isolates from urinary tract, blood, and wound infections as well as other bodily fluids were significantly associated with the prevalence of ESBLs (). One hundred percent and 33.6 % of E. coli isolates from wound and urinary tract infections were ESBLs-containing, respectively (P < 0.05). ESBL-producing isolates were determined using the phenotypic disk technique in which 53 (41.7 %) isolates were found to produce ESBL enzymes. After PCR detection, all phenotypically-positive isolates contained ESBL genes. The bla-TEM gene was the major ESBL type observed in 86.7 % of ESBL-producing isolates. Among 53 identified ESBL-producing E. coli, 81.1 % and 5.6 % were bla-CTXM and bla-SHV, respectively. However, the majority of ESBL-producing isolates were susceptible to imipenem (94.4 %) and amikacin (90.6 %).
Antibiotic susceptibility results also demonstrated that antibiotic resistance patterns of ESBL-producing E. coli are different from non-ESBL ones. ESBL-positive E. coli showed high level resistance (>80 %) to third-generation cephalosporins, but resistance to third-generation cephalosporins in ESBL-negative E. coli was low. Furthermore, resistance to fluoroquinolone antibiotics was higher in ESBL-positive E. coli than in ESBL-negative E. coli. In this study, a direct relationship between the presence of bla-CTXM and bla-TEM genes in E. coli and resistance to some antibiotics such as gentamicin, ciprofloxacin, ampicillin, ceftriaxone, ceftazidime, cefotaxime, imipenem, and co-trimoxazole were observed (P < 0.05). Isolates producing bla-CTXM were more resistant than those producing bla-TEM and bla-SHV. For example, resistance to ciprofloxacin was higher in bla-CTXM-producing isolates than in the others (p < 0.05).
Overall, the isolates were screened for the presence of the qnr gene by PCR; 8 (7.2 %) isolates containing this gene were observed. qnrS was the most dominant gene among the qnr genes. Of the 8 qnr-producing E. coli isolates, only one isolate produced qnrB, but qnrS was detected in all of the 8 isolates. No qnrA gene was identified in this study. After analysis, the data showed that 3 (37.5 %) qnr-producing isolates contained ESBL genes as well, but 5 (62.5 %) isolates were without these genes. The qnr-producing isolates were found in those E. coli-producing ESBLs that had both bla-TEM and bla-CTXM genes, but they were not found in bla-SHV-producing E. coli.
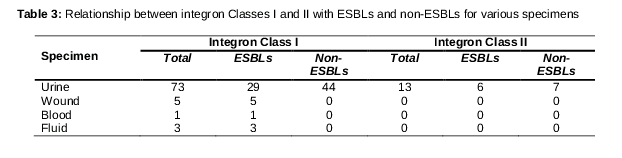
Based on data analysis, there was no significant association between ESBL and the qnr gene. Interestingly, the prevalence of the intI gene was observed in the majority of isolates such that 82 (64.5 %) isolates presented this gene, but only 13 (10.2 %) of the 127 E. coli isolates carried the intII gene. Class I integrons were more common in ESBL isolates than in non-ESBL isolates. In this study, a significant association between ESBL and integron class I was observed (P < 0.05). Conversely, no association was found between integron class II and ESBL genes.
Discussion
In the current sudy, ESBL-producing E. coli was found to be more than 40 % in the clinical samples. In addition, the results suggest the emergence of a high frequency of ESBLs inclinical isolates, which may have been caused by the excessive use of broad-spectrum antibiotics in hospitals. In fact, resistance to β-lactam antibiotics and cephalosporins was more common than to other antibiotics because of the lack of attention paid to antibiotic consumption. Ampicillin and co-trimoxazole had the highest percentages of resitance, but the highest percentages of susceptibility were observed for imipenem, amikacin, ciprofloxacin, and gentamicin. The current findings are in congruence with those of other studies conducted by Calbo et al. and Mehrgan et al [4,18].
It is noteworthy that in this study, ESBL enzymes were detected in the majority of isolates in which bla-TEM was the most common ESBL isolated from clinical samples. Furthermore, the prevalence of bla-CTXM and bla-SHV in E. coli isolates was high. demonstrates that the resistance rate of ESBL-producing isolates to all antibiotics was higher than that of non-ESBL-producing ones, especially to ampicillin, co-trimoxazole, and ceftazidime. These results are congruent with several studies conducted in Iran and other countries. According to a study done by Zaniani et al, ESBL was found in 43.9 % of isolates, demonstrating the prevalence of ESBL in E. coli isolates [19]. The results of Zaniani et al. corroborate those of the current study. Also, Calbo et al demonstrated the prevalance of ESBL in E. coli isolates and detected TEM (60 %), SHV (16 %), and CTX-M (24 %) were also detected in these bacteria [18].
Some studies in Iran have demonstrated that ESBLs have been increasing during recent years. For example, Pakzad et al and Nakhaei Moghaddam et al reported ESBL-containing E. coli in 28 % and 33 % of isolates, respectively [20,21]. Based on the results of the above-named studies, the increase in ESBL genes found in E. coli isolates in the last few years is a major problem. The current study found the prevalence rates of the intI and intII genes in E. coli isolates to be 82 (64.5 %) and 13 (10.2 %), respectively.
These results are similar to results obtained by Elisabete Machado et al [12]. Moreover, in a study by Aziz Japoni et al, the existence of integrons was confirmed in 44.8 % of E. coli isolates, and the prevalence of integrons was about half the amount obtained in the current study [22]. The prevalence of integrons was 55.5 % in Mexico and 26.1 % in Turkey [23,24]. These results confirm that the prevalence of integron genes varies between Iran and other parts of the world. This resistance can probably be associated with a combination of ESBL genes and integron class I genes.
Integrons play an important role in the antibiotic resistance of clinical E. coli strains, and they are able to capture, integrate, and express those gene cassettes encoding antibiotic resistance [12]. In this study, a significant association between ESBLs and integron class I genes was observed as it was in several other studies the results of which agree with the current results [12,25]. Therefore, it can be said that ESBLs genes are located on class I integrons.
Nevertheless, 8 (7.2 %) qnr-positive E. coli isolates were identified among a total of 127 clinical isolates. Although the prevalance of qnr genes was low, this plasmid-borne mechanism is a major concern for increasing antibiotic resistance in other isolates. There results have also been confirmed by other investigators, such as Mammeri et al. who reported that qnr genes were only found in 0.3 % of E. coli isolates [13]. Similar to the current study, Jeong et al. demonstrated that the prevalence of qnr genes in E. coli isolates was low [7].
Conclusion
The findings of this study indicate that amikacin, imipenem, ciprofloxacin, and gentamicin are the most appropriate oral antibiotics for treating ESBL infections in Kerman, Iran, but ampicillin is not a good choice for treating E. coli infections. The high prevalence of ESBL and intI genes in the hospital setting can cause antibiotic resistance. Since available treatment options are limited, antibiotic control policies combined with the implementation of infection control measures are of utmost importance. Furthermore, adherence to health recommendations in hospitals and appropriate prescribing of antibiotics can minimize antibiotic resistance resulting from ESBLs, integrons, and qnr-borne bacteria in the future.
Declarations
Acknowledgement
References
Archives


News Updates